 |
|
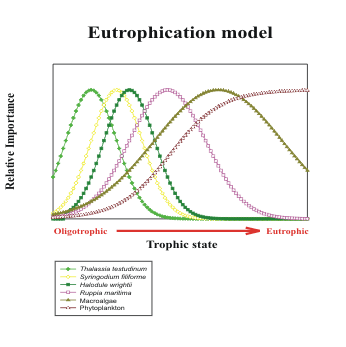
Figure 2. Idealized change in relative dominance of species along
a eutrophication gradient in the FKNMS |
Each species in the species dominance-eutrophication
gradient model (Figure 2) can potentially dominate over a
range of nutrient availability and the model predicts a change
in species dominance as nutrient availability changes. These
changes are not instantaneous, however. Field evidence
suggests that species replacements may take place on a time
scale of a decade or more. It is desirable that we be able to
predict the tendency of the system to undergo these changes
in species dominance before they occur, so that management
actions can be taken. Tissue nutrient concentrations can be
monitored to assess the relative availability of nutrients to the
plants. For phytoplankton communities, this idea is captured
in the interpretation of elemental ratios compared to the
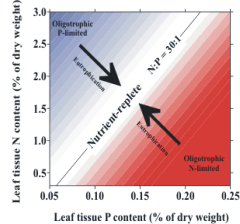
familiar "Redfield ratio" of 106C:16N:P. For the seagrass T.
testudinum, the critical ratio of N:P in green leaves that
indicates a balance in the availability of N and P is ca. 30:1,
and monitoring deviations from this ratio can be used to infer
whether N or P availabilities are limiting this species’ growth.
Hence, T. testudinum is likely to be replaced by faster-growing competitors if nutrient availability is
such that the N:P of its leaves is ca. 30:1. A change in the N:P in time to a value closer to 30:1 is
indicative of eutrophication (Figure 3).
 |
|
Figure 3. Relative amounts of nitrogen and
phosphorus in seagrass leaves change in a
predictable way as nutrient availability
increases. |
In addition to elemental content, the stable isotopic composition of plant tissues change as
environmental conditions change. As light availability to the seagrasses is reduced, as occurs when
nutrient availability increases and faster growing taxa proliferate, the stable carbon isotopic composition
shifts towards values more depleted in the heavier stable C isotope, 13C. Hence, long-term changes in
d13C values of seagrasses can serve as an indicator of changing light environment. Nitrogen stable
isotopic composition can also indicate change in nutrient status; however this indicator is much more
complicated to interpret. Sewage-derived nitrogen tends to be enriched in the heavier stable isotope,
15N, while fertilizer-derived nitrogen is depleted of this heavier isotope. Additionally, changes in the
availability of light may affect plant stable N contents in a manner similar to the stable carbon isotopic
content. While interpretation of patterns in stable N isotope composition are not straightforward,
changes in the d15N of seagrass tissues does indicate a change in the nutrient environment of those
plants.
These models lead directly to a definition of trends likely to be encountered in the seagrass communities
of south Florida if humans are causing regional changes in nutrient availability because of alterations to
quantity and quality of freshwater inputs to the marine ecosystem:
1) regional eutrophication will cause N:P ratios of seagrasses to approach 30:1 from higher or
lower values indicative of oligotrophic conditions.
2) regional eutrophication will cause a shift in species dominance in south Florida seagrass beds.
The first responses to eutrophication will be evidenced by an increase in the relative abundance
of fast-growing seagrass species (H. wrightii and S. filiforme) at the expense of the now-dominant, slow-growing T. testudinum. At later stages of eutrophication, macroalgae and
microalgae will become the dominant primary producers.
3) eutrophication will cause a decrease in the d13C of seagrass leaves.
4) a change in nutrient availability will cause shifts in the d15N of seagrass leaves.
Our monitoring data indicates a large spatial gradient in the N:P ratios of Thalassia testudinum across
the sanctuary, with N:P ratios predicting nitrogen limitation in the offshore parts of the sanctuary and
predicting phosphorus limitation in the nearshore areas. These predictions based on our conceptual
model were tested experimentally and reported on in previous fiscal year reports.
The nearshore seagrass
beds did not respond strongly to nutrient addition (see Ferdie, M. and J.W. Fourqurean, 2004.
Responses of seagrass communities to fertilization along a gradient of relative availability of nitrogen
and phosphorus in a carbonate environment. Limnology and Oceanography 49(6):2082-2094). We
also have completed experiments investigating the role of grazing fish in controlling the distribution of
seagrasses in the seagrass beds adjacent to coral reefs in the FKNMS, and we have found that
herbivorous fish can exert a greater control than nutrient availability in controlling seagrass community
composition along the reef tract of the FKNMS (see Armitage and Fourqurean, in press. Effects of
herbivory differ between seagrass species in a coral reef environment. Journal of Experimental Marine
Biology and Ecology).
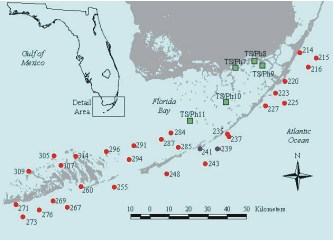
At 7 nearshore Level 1 sites in the Florida Keys, there have been changes in the relative abundance of
seagrasses and macroalgae over the period 1995 - 2005 that are consistent with increased nutrient
availability (Figure 4). At none of these has there yet been a decrease in seagrass abundance, but our
conceptual model predicts that increases in fast-growing macroalgae should precede decreases in
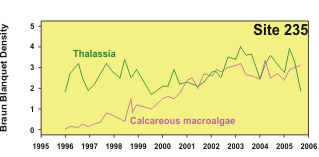
seagrass abundance (Figure 2). One example, from site 235 offshore of Lower Matecumbe Key, shows
how macroalgae have steadily increased in abundance over the monitoring period (Figure 5).
 |
 |
|
Figure 4. Red dots indicate Level 1 sites showing changes in
relative abundance of primary producers consistent with
eutrophication. |
Figure 5. An example of a site exhibiting changes in relative
abundance of benthic primary producers. See Figure 1 for
location. |
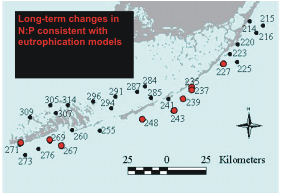
In addition to these sites where relative abundance of
primary producers has changed, at 9 of 30 Level 1 sites
there have been long-term shifts in the ratio of nitrogen
to phosphorus in seagrass leaves that are consistent
with increases in nutrient availability (Figure 6).
 |
|
Figure
6. Red dots indicate Level 1 sites showing
changes in N:P of primary producers consistent with
eutrophication |
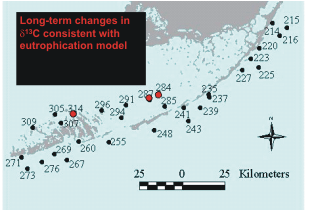
Stable isotopes of C and N of seagrasses also showed some significant long-term trends at some of the
Level 1 sites. Decreasing values of d13C, which are consistent with a decrease in light reaching the
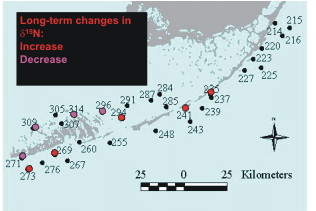
seagrasses, were found at 3 sites (Figure 7). We measured both increases and decreases in stable
nitrogen isotopic composition, with a geographic grouping of sites that increased and decreased (Figure
8).
 |
 |
|
Figure
7. Location of Level 1 stations exhibiting a decrease
in stable isotope ratios of carbon in seagrasses, consistent
with a decrease in light available to the plants. |
Figure
8. Locations of Level 1 sites showing an increase (red
symbols) of decrease (pink symbols) in stable isotope rations
of nitrogen. |
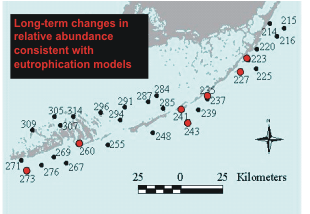
The sites that showed changes consistent with increased nutrient availability were not randomly
distributed across the Sanctuary - rather, all of these sites were relatively close to shore in the Middle
and Lower Florida Keys (Figure 9). The lack of any such changes in the Upper Florida Keys suggests
that the factor driving the observed changes is not present across the entire Sanctuary, so factors acting
at the global scale (like global warming or coastal overfishing) are not likely responsible for the observations. In addition to Level 1 sites that are exhibiting changes that are consistent with long-term
increase in nutrient supply, two additional sites were severely impacted by hurricanes over the course of
the monitoring period. While the trends in indicators we present are consistent with model predictions of
nutrient-induced changes of these systems, there may be reasons for these observations that are
unrelated to man's activities in the region. However, the spatial pattern of changes and the agreement of
the changes with models of the system suggest that there is regional-scale change in nutrient availability
that is causing changes in seagrass beds over a wide portion of the FKNMS.
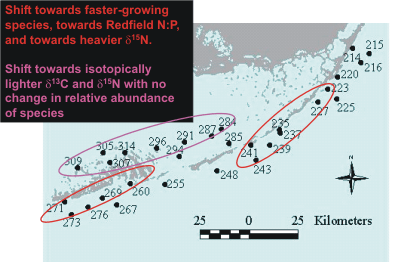 |
|
Figure
9. Locations of Level 1 sites showing long-term trends consistent
with eutrophication over the period 1995-2005, and the identity of the
changing indicators. |
In the previous 2 years of this project, resurveying the Level 2 and Level 3 sites revealed no spatially
consistent patterns in changes in relative abundance of seagrass communities from throughout the
sanctuary. However, this year we detected significant changes in the abundance and distribution of two
seagrass species (Table 1). The changes in Braun Blanquet density for Thalassia testudinum and
Syringodium filiforme for the period 1998 and 2005 were not significantly different from zero (mean
change in Braun Blanquet density = +0.08 + 0.09 for T. testudinum and +0.04 + 0.09 for S. filiforme
based on 355 pairwise comparisons), but there were significant decreases in the abundance and spatial
extent of both Halophila decipiens (mean decrease in density of 0.25 + 0.08) and Halodule wrightii
(0.06 + 0.04). The number of sites supporting H. decipiens decreased from 87 in 1998 to 26 in 2005;
and the number supporting Halodule wrightii decreased from 65 to 46. Whether these changes were
caused by environmental forcing or the ephemeral nature of these annual species is currently being
investigated. In FY 2006, we will resample an additional 250 sites that were surveyed in 1999.
|
Table 1. Results of resurveying 363 Level 2 and Level 3 sites in 2005 that had originally been surveyed
in 1998. |
|
|
1998
|
2005
|
|
|
|
Number of sites where present
|
206
|
201
|
|
1.1
|
1.2
|
|
0.3
|
0.3
|
|
0.0-4.9
|
0.0-5.0
|
|
|
|
Number of sites where present
|
128
|
120
|
|
0.6
|
0.6
|
|
0.0
|
0.0
|
|
0.0-5.0
|
0.0-5.0
|
|
|
|
Number of sites where present
|
65
|
46
|
|
0.1
|
0.1
|
|
0.0
|
0.0
|
|
0.0-4.9
|
0.0-5.0
|
|
|
|
Number of sites where present
|
87
|
26
|
|
0.29
|
0.03
|
|
0.0
|
0.0
|
|
0.0-4.3
|
0.0-1.6
|
|
Our surveys have provided clear documentation of the distribution and importance of seagrasses in the FKNMS. The seagrass bed that carpets 80% of the FKNMS is part of the largest documented contiguous seagrass bed on earth. These extensive meadows are vital for the ecological health of the FKNMS and the marine ecosystems of all of south Florida. Maps of spatial distributions and time series of species composition, seagrass productivity, nutrient availability and physical parameters can be found for each permanent monitoring site on the web site,
www.fiu.edu/~seagrass or on the CD (2002) or DVD (2003,
2004 & 2005) produced and submitted for the technical report at year end.
Detailed analyses of the monitoring data have led to 21 peer-reviewed publications in the scientific
literature (listed below). These publications address aspects of the functioning, status and trends of
benthic communities as well as lay the groundwork for forecasting future anthropogenic impacts on this
ecosystem. Numerous presentations by PI and students are also listed below.
Publications
1. Sea urchin overgrazing of a large seagrass bed in outer Florida Bay. Rose, C.D., W.C. Sharp, W.J. Kenworthy, J.H.
Hunt, W.G. Lyons, E.J. Prager, J.F. Valentine, M.O. Hall, P. Whitfield, and J.W. Fourqurean. 1999. Marine Ecology
Progress Series 190:211-222.
2. Spatial and temporal pattern in seagrass community composition and productivity in south Florida. Fourqurean, J.
W., A. Willsie, C. D. Rose and L. M. Rutten. 2001. Marine Biology 138:341-354.
3. Competition between the tropical alga, Halimeda incrassata, and the seagrass, Thalassia testudinum. Davis, B.C. and
J.W. Fourqurean. 2001. Aquatic Botany 71(3):217-232.
4. Large-scale patterns in seagrass (Thalassia testudinum) demographics in south Florida. Peterson, B. J. and J. W.
Fourqurean. 2001, Limnology and Oceanography 46(5):1077-1090.
5. Seagrass distribution in south Florida: a multi-agency coordinated monitoring program. Fourqurean, J. W., M. J.
Durako, M. O. Hall and L. N. Hefty. 2002. Pp 497-522 in: Porter, J.W. and K.G. Porter, eds. The Everglades,
Florida Bay, and the coral reefs of the Florida Keys. CRC Press LLC, Boca Raton. 1000pp.
6. Disturbance and recovery following catastrophic grazing: studies of a successional chronosequence in a seagrass
bed. Peterson, B. J., C. D. Rose, L. M. Rutten and J. W. Fourqurean. 2002. Oikos 97: 361-370.
7. Seagrass nutrient content reveals regional patterns of relative availability of nitrogen and phosphorus in the Florida
Keys, FL, USA, Fourqurean, J. W. and J. C. Zieman. 2002. Biogeochemistry 61: 229-245.
8. Monitoring of soft-bottom marine habitat on the regional scale: the competing goals of spatial and temporal
resolution. Fourqurean, J.W. and L.M. Rutten. 2003. Pp 257-288 in: Busch, D. and J.C. Trexler, eds. Ecological
Monitoring of Ecosystem Initiatives. Island Press, Washington, D.C.
9. Forecasting the response of seagrass distribution to changing water quality: statistical models from monitoring data.
Fourqurean, J.W., J.N. Boyer, M.J. Durako, L.N. Hefty, and B.J. Peterson. 2003. Ecological Applications 13(2):
474–489.
10. Intra- and interannual variability in seagrass carbon and nitrogen stable isotopes from south Florida, a preliminary
study. Anderson, W.T. and J.W. Fourqurean. 2003. Organic Geochemistry 34:185-194.
11. Spatial and seasonal variability in elemental content, δ13C and δ15N of Thalassia testudinum from south Florida.
Fourqurean, J.W. S.P. Escorcia, W.T. Anderson and J.C. Zieman. Estuaries. 2005. 28(3):447-461.
12. The impact of Hurricane Georges on soft-bottom, backreef communities: site- and species-specific effects in south
Florida seagrass beds. Fourqurean, J. W. and L. M. Rutten. Bulletin of Marine Science. 2004. 75(2):239-257.
13. Nutrient limitation of benthic primary production in the upper Florida Keys. Ferdie, M., Masters of Science Thesis,
Florida International Universtiy, Biology Department, 2002.
14. An assessment of nearshore benthic communities of the Florida Keys. Rutten, L.M., Masters of Science Thesis,
Florida International University, Biology Department, 2002.
15. Elucidating seagrass population dynamics: theory, constraints and practice. Fourqurean, J.W., N. Marbà, and C.M.
Duarte. 2003. Limnology and Oceanography 48(5):2070-2074.
16. Differential responses of benthic primary producers to nitrogen and phosphorus enrichment in a carbonate coastal
marine system. Ferdie, M. and J.W. Fourqurean. 2004. Limnology and Oceanography 49(6):2082-2094.
17. Spatiotemporal variation of the abundance of calcareous green macroalgae in the Florida Keys: A study of
synchrony within a macroalgal functional-form group. Collado-Vides, L., L.M. Rutten and J.W. Fourqurean. 2005.
Journal of Phycology 41(4):742-752.
18. Dynamics of segrass stability and change. Duarte, C.M., J.W. Fourqurean, D. Krause-Jensen, and B. Olesen. 2006.
Pp. 271-294 in Larkum, A.W.D., R.J. Orth and C.M. Duarte (eds) Seagrasses: Biology, ecology and conservation.
Springer, Dordrect.
19. Potential role of the sponge community in controlling phytoplankton blooms in Florida Bay. Peterson, B.J., C.M.
Chester, F.J. Jochem and J.W. Fourqurean. Marine Ecology Progress Series. In press.
20. The short-term influence of herbivory near patch reefs varies between seagrass species. Armitage, A.R and J.W.
Fourqurean. Journal of Experimental Marine Biology and Ecology. In press.
21. Phenology, sexual reproduction, and the factors affecting sexual reproduction of the marine angiosperm, Thalassia
testudinum, in the Florida Keys National Marine Sanctuary (FKNMS). Cunniff, K. M. Masters of Science Thesis,
Florida International University, Biology Department. 2006.
Presentations